Una tarea compleja")
El ensamblaje y la función de los cilios requieren transporte intraflagelar (IFT), un mecanismo que utiliza «trenes» para transportar cargas dentro y fuera de los cilios. Si bien se ha aprendido mucho sobre IFT en las últimas décadas, el ensamblaje de trenes IFT, la carga de mercancías y la regulación del transporte siguen siendo poco conocidos. En un estudio reciente, Hesketh, Mukhopadhyay y sus colegas obtuvieron la estructura completa del complejo IFT-A, un elemento clave de los trenes IFT. Al modelar IFT-A en trenes anterógrados y realizar mutagénesis guiada por estructuras, los autores descubren cómo el complejo IFT-A polimeriza y forma vagones para llevar a cabo sus distintas funciones.
Los cilios son orgánulos basados en microtúbulos que sobresalen de la superficie de la mayoría de las células eucariotas. Sus propiedades funcionales únicas les permiten participar en diversas funciones sensoriales, vías de señalización, flujo de fluidos y motilidad celular. Por lo tanto, no sorprende que los defectos de los cilios a menudo conduzcan a una amplia gama de trastornos humanos, denominados colectivamente ciliopatías, que afectan múltiples sistemas de órganos y tejidos.
La ciliogénesis y las funciones ciliares dependen de un sistema de transporte bidireccional conocido como transporte intraflagelar (IFT; revisado en la ref. 1). Durante la IFT anterógrada, máquinas moleculares grandes, llamadas trenes de IFT, son impulsadas por motores Kinesin-2 para transportar carga y moléculas de señal hacia la punta ciliar. Una vez en la punta, se descarga la carga, se reacondicionan los trenes y se activan los motores Dynein-2 para impulsar el IFT retrógrado hacia la base ciliar.
Dos de los componentes principales de los trenes IFT son los complejos IFT-A e IFT-B, compuestos por 6 y 16 subunidades, respectivamente.1. El complejo IFT-B es fundamental para iniciar la IFT anterógrada, ya que sirve como andamio sobre el que se monta el tren IFT.dos. Por otro lado, los mutantes del complejo IFT-A generalmente conducen al desarrollo de pestañas cortas y voluminosas, con grandes acumulaciones de componentes IFT en sus puntas, lo que indica que IFT-A es importante para IFT retrógrada. Curiosamente, también se ha demostrado que el complejo IFT-A desempeña un papel crucial en la importación de proteínas de membrana a los cilios (revisado en la ref. 1). No está claro si estas funciones aparentemente distintas del IFT-A están relacionadas entre sí. Además, aunque la organización general del complejo IFT-A se ha estudiado hasta cierto punto, su montaje y modo de operación en el tren IFT siguen siendo difíciles de alcanzar.
En un estudio elegante y completo, Hesketh, Mukhopadhyay y colegas3 reconstituyó el complejo IFT-A humano in vitro y obtuvo su estructura en microscopía crioelectrónica (crio-EM) con una resolución que va de 3,5 a 15 Å. Como se esperaba de estudios bioquímicos previos1, los autores muestran que la estructura del complejo IFT-A se puede subdividir en dos módulos (Fig. 1). Descubrieron que el módulo anteriormente conocido como núcleo IFT-A en realidad está compuesto por IFT144, IFT140 y la mitad del terminal C de IFT122. Curiosamente, este módulo resulta ser la región más flexible del complejo IFT-A. Por otro lado, las subunidades IFT-A no centrales, IFT121, IFT139 e IFT43, junto con la mitad N-terminal de IFT122, constituyen en realidad el módulo más rígido de este complejo proteico. Pensando en esta nueva organización estructural, los autores renombraron estos módulos como IFT-A1 e IFT-A2, respectivamente.3.
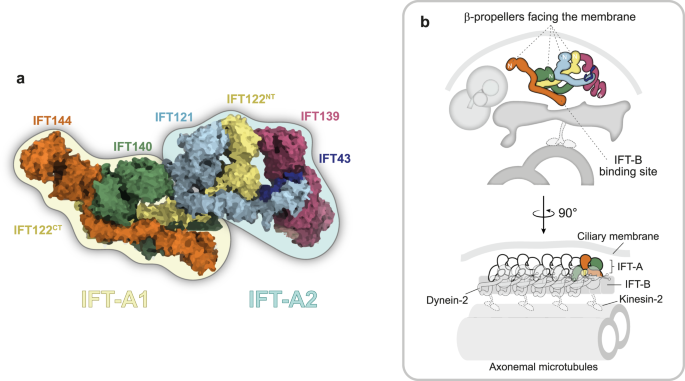
un El complejo IFT-A se subdividió en dos módulos: IFT-A1 (resaltado en amarillo claro) e IFT-A2 (resaltado en azul claro). IFT-A1 abarca IFT144, IFT140 y la región C-terminal (CT) de IFT122, mientras que IFT-A2 se compone de IFT121, IFT139, IFT43 y el N-terminal (NT) de IFT122. B Ilustración de IFT-A en vistas ortogonales en relación con IFT-B, axonema, membrana ciliar, dineína-2 y quinesina-2 (posición inferida, por contexto; no a escala). La región N-terminal de cada subunidad IFT-A se indica con una N en blanco. Arte por: María JG De-Castro.
Extendiendo las interacciones establecidas en los módulos IFT-A, los autores encontraron que el ensamblaje del complejo IFT-A está impulsado principalmente por la dimerización de dos conjuntos de subunidades: IFT144 con IFT140 e IFT121 con IFT122. Sorprendentemente, ambas interacciones están mediadas por regiones TPR en estas subunidades, todas las cuales contienen una hélice α característica entre el segundo y el tercer TPR. De acuerdo con la importancia de este dominio distinto en el ensamblaje de IFT-A, los autores encontraron que la mutación de los residuos de interfaz clave en IFT140 TPR interrumpe casi por completo su unión a IFT144. Además, la expresión de este mutante IFT140 en células IMCD3 no logra rescatar los defectos de ensamblaje del cilio asociados con la pérdida de IFT140, lo que subraya la relevancia funcional de esta interacción.3.
A continuación, para obtener información sobre la polimerización IFT-A, los autores ajustaron su modelo atómico del complejo IFT-A al mapa de tomografía crioelectrónica (crio-ET) de los trenes IFT anterógrados de Chlamydomonas reinhardtii (EMDB: 4304), obtenido previamente por el laboratorio Pigino4. Después de que el módulo IFT-A2 más suave se ajustara inequívocamente a la media del subtomograma IFT-A, el módulo IFT-A1 más suave se ajustó bien a las densidades restantes utilizando el ajuste de dinámica molecular flexible. En particular, el IFT122 juega un papel clave en esta configuración, ya que sirve como puente flexible que conecta los dos módulos IFT-A, facilitando los cambios conformacionales necesarios para la polimerización IFT-A en trenes IFT.3 (Figura 1).
Esta asignación de subunidades IFT-A en el mapa crio-ET permitió a los autores investigar también los vínculos establecidos entre los complejos IFT-A e IFT-B en trenes anterógrados. Un estudio anterior ya había identificado una interacción entre IFT144 y la subunidad IFT-B IFT885. De acuerdo con esto, los autores observaron que la región C-terminal de IFT144 se coloca en estrecho contacto con el complejo IFT-B (Fig. 1). Además, utilizando AlphaFold, los autores mapearon con éxito el motivo de interacción de IFT144 con IFT88 y descubrieron que esta interacción se ve facilitada por un conector flexible en el terminal C de IFT88. La mutación de residuos conservados en el motivo de unión a IFT88 de IFT144 redujo en gran medida la carga de IFT144 en los cilios, presumiblemente dejando el complejo IFT-A completo varado en la base ciliar. La importancia del vínculo establecido entre estos componentes de IFT-A e IFT-B se enfatiza aún más por el hecho de que esta forma mutante de IFT144 no pudo restaurar la elongación del cilio en las células inactivadas de IFT144.3.
Posteriormente, los autores alinearon múltiples copias del complejo IFT-A en el mapa crio-ET de trenes anterógrados para crear un modelo molecular del polímero desde un punto de vista más biológico. Al atribuir los diferentes dominios de las subunidades IFT-A a las diversas densidades en el polímero, observaron que los impulsores β de IFT144, IFT140, IFT122 e IFT121 se dirigen a la membrana ciliar (Fig. 1), de acuerdo con un papel en la importación de proteínas de membrana para los cilios. Curiosamente, los complejos IFT-A adyacentes establecen múltiples puntos de conexión entre ellos, tanto a nivel de controladores β como de dominios TPR. Esta disposición entrelazada conduce a la formación de compartimentos vacíos cerca de la membrana ciliar, que los autores denominaron «carros» IFT.3.
A continuación, los autores muestran cómo los transportadores IFT-A se asocian con las proteínas adaptadoras TULP, los principales mediadores de la importación de proteínas de membrana a los cilios. Usando AlphaFold, identificaron un canal dentro de IFT122 CT como el sitio principal para la unión a la proteína TULP. De hecho, la mutación de los principales residuos de la interfaz en esta región de IFT122 suprime la interacción IFT-A/TULP sin alterar la integridad del complejo IFT-A. Importante, expresando este IFT122TULPmut falla en el rescate de la incorporación de la proteína de la membrana de los cilios en células knockout para IFT1223similar a lo que se había observado en el pasado con la pérdida de adaptadores TULP1.
Tras un examen más detallado, los autores también encontraron que los elementos clave de la maquinaria IFT, a saber, IFT-A, IFT-B y el motor retrógrado Dynein-2, se acumularon en las puntas de los cilios que expresan IFT122TULPmut. Estos hallazgos sugieren que la presencia del complejo IFT-A por sí sola no es suficiente para activar la IFT retrógrada y recuperar estos componentes en la base de los cilios. Sorprendentemente, encontraron que ICK, una quinasa ciliar previamente implicada en la modulación de IFT6estuvo ausente de IFT122TULPmut pestañas3. Este emocionante resultado reveló que IFT-A controla la localización ciliar de ICK, lo que permite a los autores establecer una conexión entre los roles de IFT-A en la importación de proteínas y en IFT retrógrada.3.
En resumen, este estudio fundamental del laboratorio de Roberts remodela nuestra visión de la estructura de IFT-A, al tiempo que proporciona información fundamental sobre cómo interactúa IFT-A con IFT-B y se polimeriza en transportadores IFT-A. Además, sienta las bases estructurales para la asociación de IFT-A con adaptadores TULP y revela su control sobre la localización de ICK.3. Finalmente, este estudio mejora nuestra comprensión actual de cómo las mutaciones en los componentes ICK, Dynein-2 e IFT-A conducen a ciliopatías graves con características superpuestas.




